
- سلولی و مولکولی
- 01. دوره جامع آزمایشگاه ژنتیک
- 02. دوره جامع کارآموزی مولکولی
- 03. دوره کارآموزی ارشد آزمایشگر
- 47. دوره آنالیز داده های NGS
- 05. دوره کارآموزی جامع کاربری دستگاه فلوسایتومتری
- 16. دوره طراحی پرایمر و اصول PCR
- 11. دوره کاربری دستگاه Real time PCR
- 46. دوره پژوهشگر شو
- 21. کارگاه QF-PCR
- 04.دوره کارآموزی جامع تکنسین آزمایشگاه ژنتیک (کاریوتایپ)
- 13. دوره جامع ارشد مهندسی ژنتیک (کلونینگ) و دوره جامع کشت سلول (رده سلول سرطانی)
- 08. دوره کشت سلول (رده سلول سرطانی)
- 15. کارگاه SDS-PAGE و وسترن بلات
- 17. دوره آنالیز کروموزوم های انسانی (مقدماتی و پیشرفته)
- 31. کارگاه معرفی تکنولوژی های ویرایش ژنوم یوکاریوتی با تاکید بر تکنولوژی مدرن CRISPR/Cas9
- میکروبیولوژی
- 09. دوره جامع ارشد مهندسی ژنتیک (کلونینگ)
- 20. کارگاه بیان، استخراج و تخلیص پروتئین از میزبان باکتریایی
- 23. نرم افزار های مولکولی
- 55. دوره کاربری دستگاه فرمانتور
- 56. دوره میکروب شناسی آزمایشگاه
- 57. ارزیابی بیان پروتئین نوترکیب
- 58. کارگاه تولید پروتئین تک سلولی
- 59. کارگاه آنالیز متابولیت های ثانویه در گیاهان دارویی
- نانو فناوری
- 45. دوره الکتروریسی
- 34. تولید نانو ذرات به روش میکروامولسیون
- 35. تولید نانو ذرات به روش آسیاب گلوله ای
- 36. تولید نانو ذرات به روش سل ژل
- 37. تولید نانو ذرات به روش هیدروترمال
- 38. متصدی تولید نانو لوله های کربنی با روش CVD
- 39. متصدی توليد نانو كامپوزيت های پايه پليمری گرما نرم
- 60. آزمایشگر ارشد کروماتوگرافی گازی
- زیست پزشکی
- 06. دوره مهندسی بافت
- 14. دوره ایمونوتراپی سرطان
- 18. کارگاه حیوانات آزمایشگاهی (موش و رات)
- 19. کارگاه آنالیز داده های زیستی
- 48. کارگاه کاربری دستگاه الایزا
- 49. کارگاه جامع ساخت هیدروژل ها (تمام عملی)
- 50. کارگاه مهندسی حاملهای دارو رسان
- 51. کارگاه جامع زیستسازگاری و آزمونهای بیولوژیک
- 64. دوره ایمونوهیستوشیمی
- 65. دوره تکنیک های نوین علوم زیستی
- 66. کارگاه واکسن های نوترکیب
- آموزشی پژوهشی
- دوره های عمومی
- 07. دوره جامع کارآموزی در آزمایشگاه (پذیرش، نمونه گیری، تزریقات با سرنگ و ونوجکت، نسخه خوانی)
- 10. آموزش کنترل کیفی، استانداردسازی و مستندسازی آزمایشگاههای تشخیص پزشکی
- 18. کارگاه حیوانات آزمایشگاهی (موش و رات)
- 48. کارگاه کاربری دستگاه الایزا
- 32. دوره آنلاین WGCNA
- 61. دوره کاربر پایگاه های علوم زیستی
- 63. تحلیلگر آزمايشات علوم زيستی و بیوتکنولوژی با نرم افزار Minitab
- برنامه نویسی
- 25. دوره کامل مجازی آموزش برنامه نویسی پایتون (مقدماتی تا پیشرفته)
- 26. دوره آفلاین جامع برنامه نویسی R (مقدماتی و پیشرفته)
- 27. دوره آفلاین جامع برنامه نویسی پیشرفته R سطح ۱(TCGA)
- 28. دوره آفلاین برنامه نویسی پیشرفته R سطح 2: functional Enrichmet و نمودارها در R
- 29. دوره آفلاین برنامه نویسی پیشرفته R (دوره ی پیشرفته ی 3: GEO)
- 30. دوره آفلاین ceRNA

طراحی پرایمر


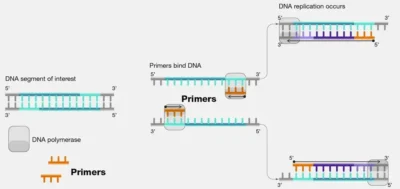
پرایمر، یک مولکول کوچک تک-رشتهای از جنس DNA است که در واکنش PCR شرکت میکند و سر 3′ هیدروکسیل آزاد را در اختیار «آنزیم Taq پلیمراز» قرار میدهد. میدانیم که در ژنوم موجودات پیچیدهای مانند انسان، در هر ناحیه، تنها یکی از دو رشته DNA به عنوان ژن فعال، عمل میکند. معمولا هدف از انجام PCR، تکثیر این ناحیه است و به آن «ناحیه هدف» (Target Region) میگوییم. برای این کار به دو پرایمر نیاز است که یکی به رشته هدف و دیگری به رشته مکمل آن متصل میشود.پرایمر، گروه هیدروکسیل آزاد در اختیار آنزیم پلیمراز قرار میدهد.
این دو پرایمر، به ترتیب، «پیشرو» (Forward) و «معکوس» (Reverse) نام دارند و با حروف اختصاری F و R نشان داده میشوند. اتصال پرایمرها به مولکول الگو بر پایه پیوندهای هیدروژنی استوار است و در هر رشته، سر 5′ پرایمر به سر 3′ مولکول الگو متصل میشود.
اتصال پرایمر
گفتیم در هر ناحیه، تنها یکی از دو رشته DNA حاوی ژن فعال است. این رشته را «آنتیسنس» (Anti-Sense) مینامیم و توالی آن، در واقع، معکوس آن چیزی است که به عنوان توالی ژن، در پایگاههای اطلاعاتی ثبت شده است.
طبق یک قرارداد جهانی، همه پایگاههای اطلاعاتی، توالی رونوشت ژن – و نه خود ژن – را ارائه میکنند. به همین دلیل است که در همه این پایگاهها، کدون آغاز تمامی توالیهای رمزکننده پروتئین، «ATG» است که مستقیما به متیونین ترجمه میشود. این توالی، در واقع همان «AUG» است که در «mRNA» وجود دارد و از روی مکمل خود (یعنی TAC) در DNA آنتیسنس رونویسی شده است.
رشته دیگر DNA که در ناحیه مورد نظر، هیچ توالی رمزکنندهای در آن دیده نمیشود، «سِنس» (Sense) نام دارد و از آن جهت به این نام خوانده میشود که توالی آن از جهت 5′→3′ دقیقا مطابق رونوشت (mRNA) است. پرایمر پیشرو، میبایست به رشته آنتیسنس متصل شود. بنابراین به گونهای طراحی میشود که نوکلئوتید انتهای 5′5′ آن مکمل نوکلئوتید مربوط، در انتهای 3′ ناحیه هدف در رشته آنتیسنس باشد.
به این ترتیب، اگر بر روی رشتههای پرایمر و رونوشت – که توالی آن به عنوان ژن، پذیرفته شده است – در جهت 5′→3′ حرکت کنیم، با ترتیب یکسانی از نوکلئوتیدها برخورد خواهیم کرد. به همین دلیل، این پرایمر را «پیشرو» (Forward) مینامیم. همین قضیه در مورد پرایمر معکوس نیز صادق است. این پرایمر به رشته DNA مقابل (رشته سِنس) متصل میشود و توالی آن از جهت 5′→3′، عکس و مکمل رشته هدف (ژن) است.
رشتههای سنس و آنتی-سنس. mRNA از روی رشته آنتی-سنس رونویسی میشود و توالی آن دقیقا مشابه رشته سنس است.

طراحی پرایمر
پیش از آنکه به بررسی اصول طراحی پرایمرها بپردازیم، میبایست با دو اصطلاح علمی آشنا شویم.
- «کارایی» (Efficiency). این اصطلاح، بیان میکند که پرایمر، تا چه حد در اتصال به توالی مکمل خود در ناحیه هدف و ایجاد محصول نهایی، موفق بوده است. کارایی بالای یک جفت پرایمر، منجر به ایجاد مقدار قابل توجهی از محصول و در نتیجه وجود یک باند بسیار قوی در ژل الکتروفورز خواهد شد.
- «اختصاصیت» (Specificity): این اصطلاح، بیان میکند که آنزیم، تا چه حد، میتواند فقط به توالی مکمل خود در ناحیه هدف، و نه هیچ نقطه دیگری از رشته الگو متصل شود. اختصاصیت بیشتر یک جفت پرایمر، به معنی کاهش باندهای غیراختصاصی در ژل الکتروفورز است.
گاهی حفظ این دو ویژگی در یک جفت پرایمر، با دشواریهایی همراه میشود به طوریکه در بیشتر موارد، افزایش یکی از این دو نیازمند کاهش دیگری است. از سوی دیگر در طراحی پرایمر، میبایست، نکاتی مانند دمای آنیلینگ، درصد GC و مواردی از این دست نیز در نظر گرفته شوند که ترکیب این موارد، فرایند طراحی پرایمر را به یک مبحث پیچیده و در عین حال کلیدی تبدیل میکند.

ویژگیهای موثر بر روی کارایی و اختصاصیت پرایمر عبارتند از:
- طول پرایمر
- «دمای ذوب پرایمر» (Melting Temperature)
- «دمای اتصال پرایمر به الگو» (Primer Annealing Temperature)
- درصد GC در طول توالی پرایمر (GC-Content)
- گیره GC (یا GC Clamp)
- ساختارهای ثانویه (Secondary Structures)
- توالیهای تکراری (Repeats)
- وجود تکرارهای پشت سر هم از یک نوکلئوتید (Run)
- پایداری سر 3′
- جلوگیری از تشکیل ساختار ثانویه در مولکول الگو
طول پرایمر
معمولا اختصاصیت یک پرایمر با تغییر در طول این مولکول، تنظیم میشود که به نوبه خود، منجر به تغییر دمای مرحله آنیلینیگ خواهد شد. در بیشتر موارد، پرایمرهایی به طول 18 تا 24 نوکلئوتید، از اختصاصیت بالایی برخوردارند و در این بازه، افزایش هر نوکلئوتید، موجب تقویت ۴ برابری اختصاصیت پرایمر میشود. کاهش طول این رشتهها به شدت بر توانایی اتصال اختصاصی آنها اثرگذار خواهد بود، به طوریکه پرایمرهایی با طول کمتر از 15 نوکلئوتید، عملا به هر نقطه از ژنوم متصل میشوند.
به طور کلی، پرایمرهایی با طول 28 تا 35 نوکلئوتید، تنها زمانی استفاده میشوند که نمونه الگو، سطوحی از «ناهمگنی» (Heterogenicity) را نشان دهد، یا پژوهشگر بخواهد یک دنباله خاص – مانند یک جایگاه برش آنزیمی – را به آن متصل کند. روشن است که در این حالت، بخشی از توالی پرایمر، در دور اول PCR به الگو متصل نخواهد شد. برای حل این مشکل، دمای آنیلینگ را در دورهای اول، پایینتر در نظر میگیرند و سپس، در دورهای بعد، آن را تا حد مجاز افزایش میدهند.
Tmپرایمر
«دمای ذوب» (Melting Temperature |Tm) یک پرایمر، دمایی است که در آن، نیمی از مولکولهای دورشتهای به حالت تک-رشتهای درمیآیند. این ویژگی، بازتابی از پایداری مولکولهای دورشتهای است. برای پرایمرهایی با طول 20 نوکلئوتید و کمتر، این دما از فرمول زیر به دست میآید:
Tm=4 (G+C)+2 (A+T)Tm=4 (G+C)+2 (A+T)
که در آن، حروف A و T و C و G، معرف تعداد نوکلئوتیدهای حاوی این بازها در رشته پرایمر هستند.
برای تخمین Tm در پرایمرهای بلندتر، از «تئوری ترمودینامیک نزدیکترین همسایه» (Nearest Neighbor Thermodynamic Theory) استفاده میشود که کمی پیچیدهتر از فرمول بالاست و با استفاده از نرمافزارهای طراحی پرایمر، محاسبه میشود. شواهد حاکی از آن است که معمولا، پرایمرهایی با Tm 52 تا 58 درجه سانتیگراد، نتایج بهتری را در PCR نشان میدهند.
دمای اتصال پرایمرها
این پارامتر را با Ta نشان میدهند و کاملا تحت تاثیر Tm پرایمرها و محصول قرار دارد. بالا بودن بیش از حد دمای Ta به معنی ناکارامدی هیبریداسیون پرایمر-الگو است که به نوبه خود، منجر به ایجاد یک محصول ضعیف در پایان PCR خواهد شد. از طرفی، پایین بودن بیش از حد این دما نیز، به تولید تعداد زیادی باندهای غیراختصاصی، میانجامد. این باندها در اثر اتصال غیراختصاصی پرایمر به الگو ایجاد شدهاند. دمای Ta از فرمول زیر به دست میآید.
Ta=0.3×Tm(Primer)+0.7×Tm(Product)−14.9Ta=0.3×Tm(Primer)+0.7×Tm(Product)−14.9
که در آن:
- Tm پرایمر: دمای ذوب ناپایدارترین دوپلکس پرایمر- الگو
- Tm محصول: دمای ذوب محصول PCR
همانطور که از این فرمول پیداست، دمای آنیلینگ، همواره پایینتر از دمای ذوب خواهد بود. در آزمایشگاه، معمولا دمای آنیلینگ، ۲ تا ۴ درجه سانتیگراد، کمتر از دمای ذوب پرایمرها در نظر گرفته میشود.
درصد GC
فراوانی نسبی نوکلئوتیدهای حاوی گوانین (G) و سیتوزین (C) که به صورت درصد بیان میشود، بر روی برخی از ویژگیهای پرایمر، از جمله Tm تاثیرگذار است. معمولا پیشنهاد میشود درصد GC پرایمرها در بازه 40 تا 60 درصد حفظ شود.
گیره GC
وجود نوکلئوتیدهای G و C در بین ۵ نوکلئوتید انتهای 3′ پرایمر، با ایجاد پیوندهای شیمیایی پایدارتر بین آن و الگو، به بهبود «کارامدی پرایمر» کمک میکند. به این ترتیب، انتهای 3′ پرایمر، مانند یک گیره محکم عمل میکند و آن را به رشته الگو متصل نگه میدارد تا آنزیم پلیمراز، کار ساخت زنجیره جدید را آغاز کند. با این وجود، همواره توصیه میشود تعداد این نوکلئوتیدها در انتهای 3′ پرایمر، بیش از ۳ عدد نباشد. چرا که در غیر اینصورت، زمینه را برای ایجاد مشکلاتی مانند تشکیل دیمرهای پایدار در سر 3′فراهم خواهد کرد.
ساختارهای ثانویه پرایمر
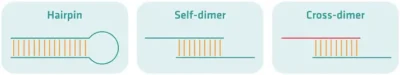
ساختارهای ثانویه در اثر برهمکنشهای بین مولکولی و درون مولکولی ایجاد میشوند و یک یا هر دو پرایمر را از دسترس آنزیم پلیمراز خارج میکنند. این مسئله موجب کاهش چشمگیر میزان محصول خواهد شد. این ساختارها در یکی از گروههای زیر قرار میگیرند:
«سنجاق سرها» (Hairpins): ساختارهای سنجاقسری در نتیجه برهمکنشهای درون-مولکولی شکل میگیرند.
خود-دیمر» (Self-Dimer): این ساختارها در اثر برهمکنش بین دو پرایمر از یک نوع رخ میدهند. همانطور که میدانیم، فراوانی پرایمرها در مخلوط واکنش، بسیار بیشتر از ژن هدف است. در این حالت، اگر مثلا پرایمر F طوری طراحی شود که بخشی از توالی آن، مکمل بخش دیگر باشد، در مرحله آنیلینگ، به جای اتصال به مولکول الگو، به پرایمر همتای دیگر متصل خواهد شد و آنزیم نیز، رشته جدیدی را از روی آن سنتز خواهد کرد.
نتیجه این امر، خارج شدن پرایمر از دسترس الگو است که به ضعیف شدن باند محصول در ژل الکتروفورز منجر میشود. در عوض، باندهای کوتاهی در پایین ژل الکتروفورز دیده میشوند که در نتیجه یک تکثیر اشتباهی ایجاد شدهاند.
دگر-دیمر» (Cross-Dimer): این ساختارها در اثر برهمکنش بین یک پرایمر F و یک پرایمر R شکل میگیرند و پیامدهایی مانند آنچه در مورد خود-دیمرها گفتیم، در پی خواهند داشت.
دیمر-پرایمرها به صورت باندهای ضعیفی در پایین ژل دیده میشوند و باند محصول را نیز تضعیف میکنند.
توالی های تکراری
منظور از این اصطلاح، وجود یک دی-نوکلئوتید تکراری در طول پرایمر است که مانند توالی «ATATAT» به صورت پشت سرهم تکرار شده است. توالیهایی از این دست، احتمال تشکیل خود-دیمرها را به شدت افزایش میدهند. یک پرایمر، حداکثر مجاز به حمل 4 دی-نوکلئوتید تکراری است.
تکرارها
منظور از این اصطلاح، وجود تکرارهای طولانی از یک نوکلئوتید در طول پرایمر است که ترادفی مانند «ACCCCCCGT» را ایجاد میکنند. چنین تکرارهایی، احتمال اتصال اشتباه پرایمر را افزایش میدهند. یک پرایمر، حداکثر مجاز به حمل 4 نوکلئوتید تکراری پشت سر هم است.
پایداری 3′
این ویژگی، بازتابی از مقدار عددی △G برای اتصال 5 نوکلئوتید انتهای 3′3 پرایمر به الگو است. ناپایداری پیوندهای این بخش، منجر به تشکیل باندهای غیر اختصاصی خواهد شد.
ساختارهای ثانویه در مولکول الگو
میدانیم که مولکولهای اسید نوکلئیکی تک رشتهای، بسیار ناپایدارند و با پیچ و تاب خوردن روی خود و تشکیل پیوندهای هیدروژنی بین بخشهای مختلف، انواع ساختارهای ثانویه را ایجاد میکنند. پایداری این ساختارها، به نوبه خود، به «اختلاف میزان انرژی آزاد» (△G) و Tm آنها وابسته است. توجه به Tm مولکول الگو، در طراحی پرایمر انواع واکنشهای PCR، به ویژه «ریل-تایم» (qPCR) از اهمیت بالایی برخوردار است.
اگر هر یک از مولکولهای الگو و پرایمرها ساختارهای ثانویهای را تشکیل دهند که حتی در دماهای بالاتر از دمای آنیلینگ نیز، از هم باز نشوند، این مولکولها قادر به تشکیل پیوند با یکدیگر نخواهند بود. به این ترتیب، میزان محصول به شدت کاهش مییابد و این امر، به ویژه در مورد qPCR – که برای تعیین دقیق و کمی میزان رونوشت یک ژن به کار میرود – موجب خطاهای آشکاری در نتیجه نهایی خواهد شد.
برای حل این مشکل، میبایست، تا حد امکان، پرایمرها برای مناطقی طراحی شوند که احتمال تشکیل ساختارهای ثانویه در آنها ضعیف یا با △G بالا باشد.
همولوژی (شباهت)
برای بهبود ویژگی «اختصاصیت» پرایمر، باید تا جایی که ممکن است، نقاطی برای اتصال آنها در نظر گرفته شوند که همولوگ بخشهای دیگری از ژنوم یا ترانسکریپتوم نباشند. در غیر اینصورت، پرایمر، علاوه بر ناحیه هدف، در تکثیر نواحی دیگر نیز شرکت خواهد کرد. این پدیده، «اتصال اشتباه» (False Priming) نام دارد و نتیجه آن، ضعیف شدن باند مورد نظر و دیدن باندهای غیراختصاصی در ژل الکتروفورز است. برای حل این مشکل، معمولا از نرمافزار BLAST استفاده میشود. این نرمافزار به صورت رایگان در پایگاه دادههای NCBI موجود است.
نرم افزارهای کاربردی طراحی پرایمر
امروزه، نرمافزارهای گوناگونی برای طراحی پرایمر، در دسترس هستند که استفاده از آنها میتواند موجب کاهش چشمگیری در وقت، هزینه و نیروی انسانی شود. نکته قابل توجه در مورد کار با این نرم افزارها، اختلاف در نتیجه خروجی آنها است. این امر، به ویژه برای پژوهشگران کمتجربهتر، ممکن است تا حدی گیجکننده باشد.
توجه داشته باشید که این اختلاف، در بیشتر موارد، ناشی از الگوریتمی است که این برنامهها با آن کار میکنند و گاهی، با تغییر برخی از پیشفرضهای برنامه، به نتایج یکسانی دست خواهید یافت. چند مورد از نرمافزارهای پرکاربرد در زمینه طراحی پرایمر در ادامه آورده شدهاند.
- «Oligo» یکی از پرکابردترین و کاربرپسندترین نرمافزارهای طراحی پرایمر به شمار میرود. این برنامه، به پژوهشگر امکان میدهد تا یک جفت پرایمر یا پروب، یا مجموعهای از آنها را طراحی، آنالیز و با هم مقایسه کند. این آنالیزها شامل بررسی ویژگیهای مختلف پرایمرهای پیشنهادی، از جمله ساختارهای ثانویه، جایگاههای اتصال اشتباهی (False Priming Sites)، پایداریهای داخلی و برخی ویژگیهای فیزیکی است. با اتصال به اینترنت، اولیگو میتواند برخی از دادههای مورد نیاز خود را مستقیما از پایگاه NCBI دریافت و ذخیره کند.
- «Primer Premier» به طور خودکار، کلیه دستورالعملهای لازم برای طراحی یک جفت پرایمر خوب را پیادهسازی میکند. با کمک آن میتوانید برای یک توالی تکی، یا یک مجموعه «همردیفشده» (Alignment) پرایمرهای اختصاصی یا تصادفی (Degenerate) طراحی کنید. این برنامه، همچنین امکان بررسی جایگاههای برش آنزیمی، آنالیز قطعات کانتیگ و طراحی پرایمر برای تعیین توالی را فراهم میکند.
- «AlleleID» و «Beacon Designer» دو نرمافزاری هستند که برای qPCR مناسبسازی شدهاند. این دو برنامه، در طراحی پرایمر و پروبهای مورد نیاز برای شناسایی کمپلکسها به کار میروند.
- «PrimerPlex» برنامهای است که در آن میتوانید پرایمرهای مورد نیاز برای PCR چندگانه (Multiplex PCR) و مطالعات چندگانه SNP در تعیین ژنوتیپ را طراحی کنید.
- GeneRunner یک برنامه حرفه ای است که هدف آن ارائه خدمات ملکولی به متخصصین ژنتیک و زیست شناسان میباشد و یک ابزار قابل اعتماد برای تجزیه و تحلیل ژن ها محسوب میشود.این نرم افزار نیازمند فایل های فرمت SEQ است که حاوی اطلاعات ژنتیکی هستند و سپس می توانند با استفاده از دکمه ORF (Open Reading Frame) تجزیه و تحلیل شوند، که کاربران را قادر می سازد تا داده ورودی را از نظر Nucleotide یا Amino Acids را تنظیم کنند.عملکرد “Restriction Analysis” این برنامه ، کاربران را قادر میسازد تا بتوانند محل های برش را به سادگی پیدا و بررسی کنند. علاوه بر این، امکان تجزیه و تحلیل انواع آنزیم ها را فراهم کرده است.از دیگر امکانات این برنامه میتوان به Oligo Analysis اشاره کرد. کاربران می توانند یک پرایمر از بیست تا سی نوکلئوتید را طراحی کنند و با کلیک بر روی دکمه Oligo ، شرح مفصلی از تمام ویژگی ها ساختاری پرایمر مورد نظر را بدست آورند.نرم افزار GeneRunner اطلاعات در مورد اسیدهای آمینه را در جداول به نمایش میگذارد و این داده ها نیز با عملکردهای تجزیه و تحلیل پروتئین، «سرکوبگر» و «جداول ترجمه» مورد استفاده قرار می گیرند. علاوه بر این، تمام اطلاعات مربوط به این عناصر قابل ویرایش است و کاربران می توانند به طور کامل المان های خود را تنظیم کنند.