
- سلولی و مولکولی
- 01. دوره جامع آزمایشگاه ژنتیک
- 02. دوره جامع کارآموزی مولکولی
- 03. دوره کارآموزی ارشد آزمایشگر
- 47. دوره آنالیز داده های NGS
- 05. دوره کارآموزی جامع کاربری دستگاه فلوسایتومتری
- 16. دوره طراحی پرایمر و اصول PCR
- 11. دوره کاربری دستگاه Real time PCR
- 46. دوره پژوهشگر شو
- 21. کارگاه QF-PCR
- 04.دوره کارآموزی جامع تکنسین آزمایشگاه ژنتیک (کاریوتایپ)
- 13. دوره جامع ارشد مهندسی ژنتیک (کلونینگ) و دوره جامع کشت سلول (رده سلول سرطانی)
- 08. دوره کشت سلول (رده سلول سرطانی)
- 15. کارگاه SDS-PAGE و وسترن بلات
- 17. دوره آنالیز کروموزوم های انسانی (مقدماتی و پیشرفته)
- 31. کارگاه معرفی تکنولوژی های ویرایش ژنوم یوکاریوتی با تاکید بر تکنولوژی مدرن CRISPR/Cas9
- میکروبیولوژی
- 09. دوره جامع ارشد مهندسی ژنتیک (کلونینگ)
- 20. کارگاه بیان، استخراج و تخلیص پروتئین از میزبان باکتریایی
- 23. نرم افزار های مولکولی
- 55. دوره کاربری دستگاه فرمانتور
- 56. دوره میکروب شناسی آزمایشگاه
- 57. ارزیابی بیان پروتئین نوترکیب
- 58. کارگاه تولید پروتئین تک سلولی
- 59. کارگاه آنالیز متابولیت های ثانویه در گیاهان دارویی
- نانو فناوری
- 45. دوره الکتروریسی
- 34. تولید نانو ذرات به روش میکروامولسیون
- 35. تولید نانو ذرات به روش آسیاب گلوله ای
- 36. تولید نانو ذرات به روش سل ژل
- 37. تولید نانو ذرات به روش هیدروترمال
- 38. متصدی تولید نانو لوله های کربنی با روش CVD
- 39. متصدی توليد نانو كامپوزيت های پايه پليمری گرما نرم
- 60. آزمایشگر ارشد کروماتوگرافی گازی
- زیست پزشکی
- 06. دوره مهندسی بافت
- 14. دوره ایمونوتراپی سرطان
- 18. کارگاه حیوانات آزمایشگاهی (موش و رات)
- 19. کارگاه آنالیز داده های زیستی
- 48. کارگاه کاربری دستگاه الایزا
- 49. کارگاه جامع ساخت هیدروژل ها (تمام عملی)
- 50. کارگاه مهندسی حاملهای دارو رسان
- 51. کارگاه جامع زیستسازگاری و آزمونهای بیولوژیک
- 64. دوره ایمونوهیستوشیمی
- 65. دوره تکنیک های نوین علوم زیستی
- 66. کارگاه واکسن های نوترکیب
- آموزشی پژوهشی
- دوره های عمومی
- 07. دوره جامع کارآموزی در آزمایشگاه (پذیرش، نمونه گیری، تزریقات با سرنگ و ونوجکت، نسخه خوانی)
- 10. آموزش کنترل کیفی، استانداردسازی و مستندسازی آزمایشگاههای تشخیص پزشکی
- 18. کارگاه حیوانات آزمایشگاهی (موش و رات)
- 48. کارگاه کاربری دستگاه الایزا
- 32. دوره آنلاین WGCNA
- 61. دوره کاربر پایگاه های علوم زیستی
- 63. تحلیلگر آزمايشات علوم زيستی و بیوتکنولوژی با نرم افزار Minitab
- برنامه نویسی
- 25. دوره کامل مجازی آموزش برنامه نویسی پایتون (مقدماتی تا پیشرفته)
- 26. دوره آفلاین جامع برنامه نویسی R (مقدماتی و پیشرفته)
- 27. دوره آفلاین جامع برنامه نویسی پیشرفته R سطح ۱(TCGA)
- 28. دوره آفلاین برنامه نویسی پیشرفته R سطح 2: functional Enrichmet و نمودارها در R
- 29. دوره آفلاین برنامه نویسی پیشرفته R (دوره ی پیشرفته ی 3: GEO)
- 30. دوره آفلاین ceRNA

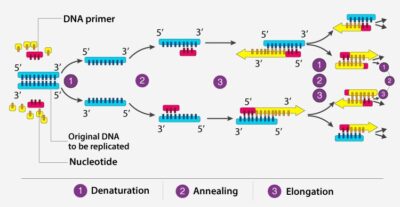
PCR و انواع آن

نگاهی کلی بر واکنش زنجیرهای پلیمراز (Polymerase Chain Reaction)
به کمک PCR میتوان ناحیه خاصی از مولکول DNA را تکثیر نمود به شرطی که توالی دو انتهای آن مشخص باشد.
سه ویژگی اصلی PCR موجب کاربردهای وسیع آن در تشخیص، تحقیقات، علوم جنایی و… گردیده است:
1) سادگی و سرعت بالا 2) حساسیت بالا که منجر به تکثیر مقادیر کم DNA حتی DNA یک سلول منفرد میشود 3) گستردگی و قدرت بالا به طوری که میتوان DNA بافتهای تجزیه شده یا فیکس شده در فرمالین را نیز تکثیر نمود.
آنزیم Taq (گرفته شده ازThermus aquaticus) فاقد فعالیت تصحیح (3´5´ اگزونوکلئازی) است اما آنزیم Pfu (گرفته شده از Pyrococcus furiosus) قدرت تصحیح دارد. دمای مناسب اتصال با تعیین دمای ذوب (Tm) محاسبه میشود.
Tm دمایی است که در آن DNA دورشتهای با بازشدن جفتبازها به DNA تک رشتهای تبدیل میشود و از فرمول زیر محاسبه میگردد:
Tm = (4 [ G+C ]) + (2[ A+T ])
- پرایمر باید مکمل رشته الگو باشد و توالی 3´ هر پرایمر نباید با هیچ قسمتی از توالی پرایمر دیگر مکمل باشد.
- طول پرایمر باید 17 نوکلئوتید (اغلب 25 – 18) باشد. طول کوتاهتر موجب تکثیر محصولات ناخواسته و طول بلند موجب کاهش سرعت هیبرید شدن آن با DNA الگو میشود.
- دمای مناسب برای هیبریدیزاسیون پرایمر و الگو باید °C 2- 1 (یا بعضاً °C5) پائینتر از دمای Tmباشد. دو پرایمر نیز باید دارای Tm یکسان (یا با کمتر از °C 5 اختلاف) باشند. تفاوت Tm پرایمرها از محصول PCR نباید بیشتر از °C10 باشند.
- به منظور اطمینان از عدم ایجاد جفتباز درونی، توالی پرایمر باید فاقد تکرارهای معکوس یا توالیهای خود مکمل بیش از bp 3 باشد.
- میزان GC در پرایمر باید 60-40 ٪ و با توزیع یکنواختی از چهار نوکلئوتید باشد.

معایب PCR در مقایسه با کلونینگ برپایه سلول
1- طول قطعه تکثیر شونده توسط PCR نباید از kb 3 بیشتر باشد و بهترین حالت طول کمتر ازkb 1 است. تکثیر قطعات kb 20 با روشهای معمول و البته با کارایی کم و قطعات تا kb 40 با روشهای خاص PCR قابل انجام است(برای این منظور از دو نوع پلیمراز مقاوم به حرارت استفاده می شود که میزان بیهینه ای از عملکرد پلیمرازی و اگزونوکلئازی 3´5´ حاصل شود).
2-میزان تولید محصول توسط PCR اندک می باشند و در حد میکروگروم محصول نهایی در دسترس قرار خواهد گرفت(راه حل استفاده از کلونینگ بر پایه سلولی می باشد).
3- آنزیم Taq با فعالیت داکسی نوکلئوتیدیل ترانسفراز انتهایی به انتهای 3´ محصول یک نوکلئوتید dA اضافه میکند که برای رفع این مسئله
الف) میتوان از وکتورهایی نظیر pGEM-T-easy با انتهای 3´ بیرون زده dT استفاده نمود که به این روش “TA Cloning” میگویند.
ب) میتوان توسط پلیمرازهای T4 یا Pfu، dA را حذف نمود(دارای خاصیت proofreading هستند).
ج) طراحی پرایمرهایی با 10 نوکلئوتید اضافی در بخش 5´ که شامل یک جایگاه برش آنزیم محدود کننده خاص می باشند که پس از انجام واکنش و تولید محصولات بتوان با شکست توسط این آنزیم، فرایند کلون کردن را به راحتی انجام داد. در مرحله اول واکنش این بخش 10نوکلئوتیدی با توالی الگو تشکیل جفت باز نمی دهد ولی در مراحل بعدی به صورت کامل تشکیل جفت باز می دهد(5´ adds on mutagenesis)
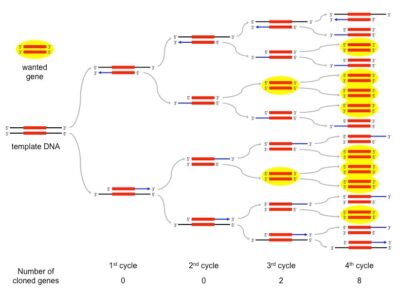
روشهای مختلف PCR و کاربرد آنها
براساس جفتشدن بازی دقیق بین انتهای3´ پرایمر و DNA الگو، تنها یک آلل خاص تکثیر مییابد. سه پرایمر، یکی برای آلل طبیعی، یکی برای آلل موتانت و دیگری به طور مشترک برای هر دو آلل نیاز است. این روش برای شناسایی جهشهای تک نوکلئوتیدی به کار میرود.
در این روش برای تکثیر توالی های اطراف یک توالی مورد شناسایی استفاده می شود که در آن از یک پرایمر مختص توالی و یک پرایمر عمومی Universal استفاده می شود. پرایمر عمومی به توالی مصنوعی که به انتهای همه توالی های هدف متصل شده است وصل می شود. در این حالت فقط یک توالی مشخص وجود دارد و فقط می توان برای آن پرایمر طراحی کرد، پس یک توالی عمومی به انتهای دیگر توالی های هدف اضافه می کنیم تا بتوانیم از یک پرایمر عمومی و یک پرایمر مکمل بخش قابل شناسایی استفاده کنیم.
(Degenerate oligonucleotide-primed) DOP – PCR کل ژنوم
برای تکثیر توالیهای مشابه درسراسر ژنوم، از پرایمرهای دژنره (مجموعهای از توالیهای اولیگونوکلئوتیدی که در برخی بخشها مشابه و در برخی بخشها متفاوت هستند) استفاده میکند.
پرایمرهای دژنره به این شکل تولید می شوندکه در حین سنتز توالی DNA در هر موقعیت به جای وارد شدن یک نوکلئوتید هر چهارنوع نوکلئوتید وارد واکنش می شوند.
مخلوط نمودن همه مواد PCR قبل از اولین مرحله دمایی دناتوراسیون، موجب افزایش شانس اتصال غیراختصاصی پرایمرها میشود. جهت کاهش این احتمال یک یا چند جزء از مواد PCR تا بعد از اولین مرحله دمایی دناتوراسیون بطور فیزیکی جدا نگهداشته میشود که این مسئله موجب افزایش اختصاصیت PCR میگردد.
روشی است برای تکثیر توالیهای نامعلوم که در مجاورت ناحیهای با توالی معلوم قرارگرفته است. ابتدا توسط آنزیم محدودگر، DNA را در اطراف توالیهای شناخته شده برش داده ، سپس محلول به شدت رقیق می شود و پس از آن به کمک لیگاز آن را حلقوی میکنند. با پرایمرهای اختصاصی توالی معلوم حلقه DNA تکثیریافته و در نهایت حلقهها را برش میدهند.
یک شکل از تکثیر غیرمتمایزاست که در آن به دو سر همه قطعات DNA حاصل از هضم با آنزیم محدودگر مولکول لینکر متصل میشود و قطعات با پرایمر اختصاصی لینکر تکثیر مییابند. لینکر و قطعات، دارای انتهای آویزان 5’ هستند تا بتوانند بههم متصل شوند.
برای تکثیر مقادیر کم DNA و افزایش اختصاصیت PCR، ابتدا ناحیهای از DNA توسط یک جفت پرایمر تکثیر میشود و از این محصول به صورت رقیق شده به عنوان توالی هدف برای دور دوم PCR توسط یک جفت پرایمر داخلی استفاده میشود.
RACE – PCR (Rapid amplification of cDNA ends):
شکلی از Anchor- primed PCR یا RT- PCR است که برای تکثیر سریع انتهای cDNA به کار میرود. در فصل یازده استراخان به صورت مفصل توضیح داده می شود.
برای افزایش اختصاصیت PCR، چرخههای دمایی را در دستگاه ترموسیکلر طوری برنامهریزی میکنند که دمای هیبریدیزاسیون از مقدار اولیه که بالای Tm مورد انتظار است بهتدریج به پائین Tm کاهش پیدا کند. درنتیجه از ایجاد محصولات ناخواسته جلوگیری میکند.
PCR چندگانه که درآن بهطور همزمان چند جایگاه روی قطعه DNA مورد نظر با چند جفت پرایمر تکثیر میشود (توجه: از این روش در تشخیص جهشهای DMD استفاده میکنند).
Alu – PCR (IRS- PCR یا IRE-PCR)
باتوجه به تعداد زیاد تکرار Alu در توالی انسان درصورتیکه دو توالی تکراری در مجاورت و با جهت مخالف هم قرار بگیرند بهوسیله یک پرایمر منفرد اختصاصی Alu میتوان توالیهای بین دو تکرار را تکثیر نمود.
تکثیر نامتمایز است که قبلاً با استفاده از پرایمرهای دژنره یا روش لینکر انجام میشد. اما در این روش ها به برخی از توالی ها تکثیر نشده باقی می ماندند به این دلیل که ساختار های ثانویه DNA برای پلیمرازهای استاندارد ایجاد مشکل می کنند و این ها از روی برخی از توالی ها می لغزند، یا اینکه از توالی ها جدا می شوند . پس محصولات تکثیر نهایی غیر اختصاصی بوده و همه ژنوم را شامل نمی شد پس برای حل این مشکل از روش تکثیر ایزوترمال غیر PCR استفاده می شود. که به آن روش (MDA; multiple displacement amplification) یا روش تغییر مکان چندگانه استفاده می شود. در این روش ازآنزیم DNA پلیمراز جایگزین کننده رشته متعلق به فاژ 29 (DNA پلیمراز جابجا کننده رشته می باشد) استفاده می شودو تکثیر به صورت حلقه چرخان می باشد، استفاده میشود. در این روش برخلاف PCR، تکثیر در دمای ثابت حدود °C 30 انجام میشود.
(Reverse transcriptase PCR) RT- PCR
دراین روش جمعیت آغازگر، RNA کل یا mRNA است که ابتدا با آنزیم ترانسکریپتاز معکوس به cDNA تبدیل شده و سپس تکثیر مییابد.

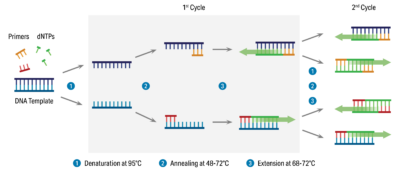
Real-time PCR
در یک آزمایش PCR توالی DNA هدف به صورت نمایی تکثیر میشود؛ به این صورت که یک الگو به دو، دو الگو به چهار و چهار الگو به هشت الگو تبدیل میشود و این روند ادامه پیدا میکند. در نگاه اول این موضوع درست به نظر میرسد. با اینحال، تکثیر نمایی نمیتواند تا ابد ادامه یابد و معمولا تا چرخه ۳۵ام سرعت واکنش کاهش خواهد یافت. در این حالت، پرایمرها و dNTPها مثل قبل به مقدار زیاد وجود ندارند؛ از بازده DNA پلیمراز کاسته شده است؛ دناتورهشدن DNAها به صورت کامل انجام نمیگیرد؛ و محصولات واکنش توسط خاصیت نوکلئازی پلیمراز از بین میروند. با وقوع موارد گفتهشده، واکنش وارد یک فاز خطی میشود. در این حالت مانند فاز لگاریتمی توالیهای الگو به طور کامل دو برابر نمیشوند. تا اینکه درنهایت تا چرخه ۴۰ام، واکنش وارد مرحله کفه (plateau) میشود که توقف تکثیر اتفاق میافتد.
پس واکنش PCR قابل تقسیم به چهار فاز کلی است: فاز اولیه، linear ground phase نام دارد. در این مرحله PCR به تازگی شروع شده است و سیگنال حاصل از سیگنال پسزمینه فراتر نرفته است. فاز دوم، exponential phase است و طی آن سیگنال فلئورسنت به طرز قابل توجهی از سیگنال پسزمینه بیشتر میشود. در این شرایط PCR در بهینهترین شرایط ممکن است و محصولات در هر چرخه دو برابر میشوند. فاز سوم Linear phase نام دارد و فاز چهارم نیز، plateau است که در آن سوبستراها کاهش مییابند و یا تخریب میشوند، آنزیم پلیمراز کارایی خود را از دست میدهد و شدت سیگنال فلوئورسنت دیگر افزایش نمییابد.
سه مرحله واکنش Real-time PCR؛ مرحله نمایی در اوایل چرخه سوم شروع میشود، با این حال دستگاه نمیتواند سیگنالهای تولیدشده در چرخههای اول را شناسایی کند. محور x نشانگر تعداد چرخههای انجامشده در آزمایش PCR و محور y نشانگر سیگنال نرمالیزهشده (Rn) است. Rn برابر نسبت شدت سیگنال رنگ reporter به شدت سیگنال حاصل از رنگ passive reference است. این رنگ معمولا ROX میباشد و در همه واکنشها حضور دارد.
Real-time PCR وابسته به توانایی دستگاه در تشخیص حداقل تعداد چرخههایی است که پس از انجامشدنشان، میزان محصولات و درنتیجه سیگنال تولیدشده به حدی باشد که بتواند از noise فلوئورسنت زمینهای دستگاه فراتر رود. این سیگنالها در محدوده تشخیص دستگاه بوده و در چرخههای اولیه متغیر هستند.
شماره چرخهای از PCR که در آن سیگنال واقعی از noise پسزمینه تشخیص داده میشود، cycle threshold و یا CT نامیده میشود. واضح است که در PCR درصورتی که شرایط برابر بوده و هیچگونه مهاری وجود نداشته باشد و تجهیزات سالم باشند، هر چه میزان نوکلئیکاسید اولیه بیشتر باشد، میزان محصولات و سرعت تشخیص آنها نیز بالاتر خواهد بود. این موضوع در مورد Real-time PCR نیز صادق است. هرچه میزان الگوهای آغازگر بیشتر باشند، سریعتر به آستانه خواهیم رسید و CT نیز کمتر خواهد شد.
این موضوع اساس سنجش مقدار DNA است. در یک واکنش Real-time PCR با بازده ۱۰۰٪ میزان محصولات در هر چرخه دو برابر میشود. به طرز مشابهی هر یک عدد تغییر در CT (از CT بیشتر به سمت CT کمتر؛ به عبارت دیگر هر یک عدد کاهش در تعداد چرخههای موردنیاز برای رسیدن به آستانه) نشاندهنده دو برابر بودن تعداد مولکولهای هدف در آغاز واکنش است. ۲ عدد کاهش نیز نشاندهنده ۴ برابر بودن تعداد مولکولهای آغازگر است. بر این اساس میتوان رابطه مقابل را نوشت: Δ CT= ۲-ΔCT fold differences
مقدار CT به صورت خودکار توسط نرمافزار دستگاه Real-time PCR تعیین میشود، اما میتواند به صورت دستی و با تعیین خط آستانه توسط اپراتور انجام گیرد. خط آستانه چه به صورت دستی رسم شود و چه به صورت اتوماتیک، محل برخوردش با منحنی تکثیر در PCR مشخصکننده CT است. CT یا چرخه آستانهای باید در قسمت نمایی نمودار و جایی که مواد اولیه واکنش فراوان هستند، باشد.
جهش زایی با PCR
5´ add-on mutagenesis: اضافه کردن یک توالی یا گروه شیمیایی ، مثلا یک جایگاه شناسایی شونده توسط آنزیم های محدود کننده، یک ریپورتر، یک گروه نشاندارمثل یک نوکلئوتید یا فلوروفر بیوتین دار، و یا یک پروموتر فاژی.
Mismatch primer mutagenesis: دو جفت پرایمر طراحی می شود ، از این دوجفت پرایمرهای ریورس یک جفت و فورواد جفت دیگر دارای جهش های منطبق بر یکدیگر می باشند. ابتدا به صورت جداگانه توسط هر یک از این جفت ها واکنش PCR انجام می شود و سپس محصولات این ها با یکدیگر مخلوط شده و با استفاده از DNA پلیمراز دو انتهی 3´هر یک از رشته های ناتمام توسط آنزیم DNAپلیمراز تکثیر شده و محصول نهایی کامل حامل جهش مورد نظر ساخته می شود.